Actualización de criterios para la suplementación de minerales traza a los bovinos
Introducción
Las células eucariotas funcionan de manera regular en un estado reducido (Veskoukis et al, 2012). Sin embargo, una cierta cantidad de especies oxidantes reactivas son necesarias para el funcionamiento normal del metabolismo; y por lo tanto son producidos de manera regular por la mayoría de los organismos; sin embargo, un desbalance en favor de los productos oxidantes desencadena una condición de peligro conocida como estrés oxidativo; bajo esta condición las estructuras de los lípidos proteínas y ácidos nucleicos pueden ser severamente alterados con el consecuente daño para los diferentes sistemas celulares y el riesgo para la viabilidad de las mismas (Seis, 1997; Birben et al., 2012; Veskoukis et al., 2012).
Los agentes oxidantes endógenos
Las especies químicas reactivas, son estructuras químicas inestables y altamente reactivas, ellos pueden ser radicales libres que poseen un número impar de electrones en su órbita más externa, pero que son capaces de tener una existencia independiente. Su vida media puede variar de unos milisegundos en los compuestos más hasta segundos y horas para los radicales más estables. Ellos pueden disparar una serie de reacciones que resulten en la oxidación de las macromoléculas (Veskoukis et al, 2012).
Las especies químicas reactivas pueden clasificarse en cuatro grupos basados en su átomo central: 1) Especies de oxigeno reactivo (ROS); 2) especies de N reactivo (NRS); 3) especies de azufre reactivas; y especies de cloro reactivas. Las más importantes de ellas son las ROS y el más abundante es el denominado “anión súper-óxido” (O2 -….), el cual es generado por la reducción de un O2 derivado de presencia de un electrón adicional (Veskoukis et al, 2012). Los principales oxidantes endógenos son el anión súper-óxido (O2 -), peróxido de hidrógeno (H2O2), radical oxhidrilo (OH-), ácido hipocloroso (HClO), radical peróxil (ROO-) y el radical hidroperóxil (HOO-), respectivamente (Birben et al., 2012).
Los ROS son producidos especialmente en la mitocondria, en donde se forman en proporción directa con la producción de energía en forma de ATP. En este organelo se produce súper- óxido y el más peligroso de los radicales el oxhidrilo (OH-), mismo que se origina por una reacción en la que participa un metal de transición, generalmente Cu o Fe (Veskoukis et al,
2012), metales en especial Fe que es necesario para el transporte de electrones de la cadena respiratoria acoplada y la fosforilación oxidativa (Huskisson et al., 2007).
El metabolismo del organismo es controlado de manera esencial por la actividad de la glándula tiroides a través de la hormonas tiroideas triyodotironina y tiroxina (Mullur et al., 2013), el adecuado funcionamiento de ambas es dependiente de la disponibilidad de yodo, lo que hace imprescindible la presencia de este mineral para el adecuado funcionamiento del metabolismo corporal.
En el mismo sentido de la producción de energía, la Vitamina B12 (Cianocobalamina), es cofactor para el funcionamiento de la metilmalonil-CoA- mutasa que participa en la síntesis de la metilmalonil-CoA, durante el proceso de oxidación ácido grasos de cadena impar y de algunos aminoácidos como Valina e Isoleucina, ambos procesos destinados a la producción de ATP, proceso que ocurre precisamente en la mitocondria; la Vitamina B12 también es imprescindible para la síntesis de los ribonucleotidos (Lenhinger, 1984; Depeint et al., 2006). En los rumiantes, de manera regular no es necesario proporcional Vitamina B12 de manera suplementaria, dado que los microorganismos del rumen son capaces de sintetizarla; tomando en cuenta que el cobalto (Co) es componente de la cianocobalamina (Huskisson et al., 2007), para que pueda ser sintetizada, es necesario que exista la disponibilidad suficiente de cobalto para la microbiota ruminal (NRC, 2000; NASEM, 2016), debido la cantidad de cobalto necesario en los rumiantes es adecuado calcularlo sobre la base de las necesidades de Vitamina B12, partiendo de este criterio, el requerimiento de cobalto para los bovinos en engorda intensiva ha sido calculado en 0.25 mg/kg de materia seca (Stangl et al., 2000).
Los sistemas antioxidantes del organismo
En los animales existen tres grupos principales de substancias con actividad antioxidante:
1) Los antioxidantes solubles en grasas: Vitamina A, Vitamina E, Carotenoides, Ubiquinonas (Coenzima Q), etcétera; 2) Los antioxidantes solubles en Agua: Ácido ascórbico (Vitamina C), Ácido Úrico, Taurina, etcétera; y 3) Las enzimas antioxidantes: Glutatión Peroxidasa, Catalasa, y Superóxido dismutasa (Surai, 2006).
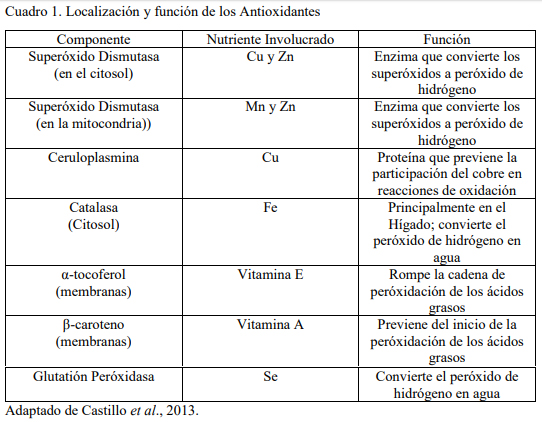
Es de mencionar, que en todos los sistemas de enzimas antioxidantes que posee el organismo animal, se requiere de la participación de algún elemento mineral para que estas puedan llevar a cabo su función (Castillo et al., 2013). En el Cuadro 1, se presenta la localización y función de una serie de antioxidantes en el organismo animal, así como los minerales y vitaminas requeridas para su funcionamiento. La superóxido dismutasa presente en el citosol, es una enzima que convierte los radicales superóxido en peróxido de hidrógeno, pero requiere para su funcionamiento de la participación de los minerales cobre (Cu) y zinc (Zn); la superóxido dismutasa presente en la mitocondria realiza la misma función y también requiere de zinc para su funcionamiento, pero no de cobre, sino de magnesio (Mg); sin embargo la ceruloplasmina que si requiere de cobre, es una proteína que previene que el cobre participe en reacciones de oxidación. La catalasa presente en el citosol y abundante en los hepatocitos convierte en peróxido de hidrógeno en agua requiere del hierro para llevar a cabo su función; y la glutatión peroxidasa necesita del selenio para orientar la reacción que lleva al peróxido de hidrógeno a transformarlo en agua.
Los minerales trazas
Se les denomina así a los minerales que son adicionados a la dieta en cantidades tan pequeñas que son expresadas en mg/kg, en los rumiantes de manera general se considera como minerales traza (o microminerales) cuyo requerimiento es conocido al cobre (Cu), cobalto (Co), Hierro (Fe), manganeso (Mn), yodo (I2), selenio (Se) y zinc (Zn). Con lo descrito anteriormente es claro que todos ellos están relacionados de manera estrecha con el metabolismo y cinco de ellos de manera muy directa con en el funcionamiento de los sistemas antioxidantes corporales (Huskisson et al., 2007; Castillo et al., 2013).
La mayor parte de las personas dedicadas a la alimentación de bovinos de engorda, utilizan como base para planear la nutrición de los animales los cuadros de requerimientos descritos en las publicaciones del Consejo Nacional de Investigación de Estados Unidos (NRC, por sus siglas en ingles). Esta prestigiosa publicación, es el producto de la reunión de un sub- comité especializado cada que se considera que el avance del conocimiento en el área requiere de una revisión de conceptos, cambios en modelos de predicción y con ello la publicación de una nueva versión del libro correspondiente. Es típico que en las publicaciones de NRC se traten los temas de energía, proteína, consumo de materia seca y agua, así como vitaminas y por supuesto minerales.
En este sentido, los requerimientos de minerales trazas de los bovinos en engorda intensiva, son frecuentemente referidos a lo que señale “el NRC”, se tiene como ejemplo el cobalto que Stangl et al. (2000) calcularon en 0.25 mg/kg de materia seca. El libro de NRC dedicado a los requerimientos de bovinos en engorda publicado hace 60 años (NRC, 1958), declara 0.10 mg de Co/kg de MS como el requerimiento de estos animales, 26 años después la publicación (NRC, 1984) señalo también 0.10 mg de Co/kg de MS, en 1996 y su actualización del 2000 (NRC, 2000) indicaron otra vez 0.10 mg de Co/kg de MS; y la más reciente publicación al respecto (NASEM, 2016) indica ¡ 0.10 mg de Co/kg de MS !
En el caso del cobre, la publicación de 1958 (NRC, 1958) señaló 10 mg /kg, a su vez en NASEM, 2016 se recomienda como el requerimiento de cobre en el alimento de los bovinos en engorda intensiva ¡10 mg /kg!; el Yodo se ha mantenido también en 0.50 mg/kg.
En relación al Hierro, el NRC (1958) mencionaba que el requerimiento debía ser menor que el de los cerdos, pero que no existían investigaciones del requerimiento de Fe en bovinos de engorda. NRC 1984 sugiere 50 mg/kg, misma cantidad encontrada en NRC, 2000 y NASEM 2016 propone ¡50 mg/kg!
Del manganeso en NRC 1958 aparece que los requerimientos son inciertos, pero que probablemente 12 mg/kg pudiera ser suficiente, en 1984 y 2000 la dosis sugerida aumentó a 40 mg/kg y NASEM 2016 se propone una disminución a 20 mg/kg.
NRC 1958 no menciona una cantidad de selenio requerida, NRC 1984 y 2000 sugieren 0.20 mg/kg y en NASEM aparece aumentado a 0.30 mg/kg
En tanto que el zinc del cual NRC 1958 declara no se conoce su requerimiento, NRC 1984 propone 30 mg/kg y tanto NRC 2000 como NASEM 2016 sostienen esa misma cantidad como el requerimiento de Zn para los bovinos en engorda intensiva. Sin lugar a dudas que a pesar de los cambios en las variables de productividad de los bovinos en engorda y los avances en tecnologías usadas en la industria, los requerimientos de minerales trazas no se hayan visto influenciados por ello.
La visón de los sub-comités para bovinos de engorda del NRC, no necesariamente reflejan lo que se ha estado haciendo en la práctica, con la incorporación de experiencia obtenida empíricamente y de resultados de investigación publicados durante ese lapso de 60 años.
A principios del siglo XXI, se publicaron los resultados de una encuesta llevada a cabo entre
24 nutricionistas que laboraban en Estados Unidos y que atendían el equivalente a 18’000,000 de bovinos en engorda intensiva (Vasconcelos y Gaylean, 2007); con las respuestas brindadas por los consultores se estableció que en promedio la cantidad de minerales trazas que ellos utilizaban en la conformación de las dietas fueron superiores a las recomendaciones del NRC; de Cobalto 0.18 mg/kg, Cobre 18 mg/kg, Yodo 0.75 mg/kg, Mn 48 mg/kg, Selenio 0.24 y Zinc 93 mg/kg. En tanto que no recomendaron la adición de Fe a la dieta al considerar que la cantidad que contienen a partir de los ingredientes primarios es suficiente para cubrir los requerimientos del animal.
Recientemente (Samuelson et al., 2016), publicaron el resultado de otra encuesta en la que participaron 24 consultores y que abarcó al equivalente de 14’000,000 de bovinos en corral. En esta ocasión los promedios de utilización de minerales traza fueron: Cobalto 0.20 mg/kg, Cobre 17 mg/kg, Yodo 73 mg/kg, Manganeso 48 mg/kg, Selenio 0.24 mg/kg y de Zinc 83 mg/kg de MS, respectivamente. Estos resultados confirman que la industria ha realizado adaptaciones en las cantidades de minerales trazas suplementados en busca de lograr un mayor productividad y de manera general los valores son muy cercanos a los de la encuesta de 2007 (Vasconcelos y Gaylean, 2007) y también por encima de las recomendaciones de NRC (2000) y NASEM (2016) que esencialmente son las mismas.
La conveniencia del uso de nivel de suplementación micro-mineral por encima de las recomendaciones de las publicaciones de NRC, ha sido frecuentemente avalada por resultados experimentales, entre ellos se encuentran los hallazgos de Stangl et al. (2000) que con criterio de utilidad en la síntesis de vitamina B12 estimaron la necesidades de cobalto en 0.25 mg/kg. Nockels et al., (1993) utilizaron una cantidad de cobre adicional de 40 mg/kg de dieta para comparar el efecto de la suplementación de este mineral a partir de fuentes inorgánicas u orgánicas en la retención de Cu en becerros. Weiss y Socha (2005), calcularon en al menos 37.8 mg/kg los requerimientos de Mn, por lo que consideraron razonable el requerimiento de Mn en 40 mg/kg de MS tanto para bovinos en engorda como para vacas lecheras que ha sido propuesto previamente (NRC, 1984; 2000). Gozzi et al. (2011) encontraron que con la adición de 0.3 mg/kg de Se a partir de levaduras enriquecidas con selenio, la ganancia de peso de novillos Charolais fue mayor y el esfuerzo al corte de la carne menor, que aquellos que recibieron la misma cantidad a de selenio a partir de selenito de sodio. Resultados similares encontraron Rossi et al. (2015). Lo que indica que el nivel de 0.3 mg de Se/kg puede ser adecuado, pero que con formas más biodisponibles o posiblemente cantidades mayores de Se pueden lograrse mayores ganancias de peso y mejor calidad de carne en los bovinos. Desde finales de los años 60’s Perry et al., (1968) comparando diferentes niveles de Zn en la dieta, encontraron que la adición del mineral en niveles por encima de los 0.30 mg/kg inducia una mejor ganancia de peso y conversión alimenticia. Barajas y Felix (2001) encontraron que con la adición de 70 mg de Zn/kg de MS (40 mg de ZnSO4 y 30 mg de Metionina de Zn) mejoró en más de 12% la ganancia de peso y en 19% la conversión alimenticia.
Autor: Dr. Ruben Brajas Cruz
Universidad Autónoma de Sinaloa (UAS México)
Literatura Citada
Barajas, R. and A. Felix. 2001. Effect of shade and organic zinc supplementation on performance of Brahman bull calves fed growing diets in dry tropic weather. J. Anim. Sci. Vol. 79 (suppl. 1):291-292.
Birben E., U. M. Sahiner, C. Sackesen. 2012. Oxidative Stress and Antioxidant Defense.
World Allergy Organization Journal, 5:9–19.
Castillo, C., V. Pereira, A. Abuelo, and J. Hernández. 2013. Effect of supplementation with antioxidants on the quality of bovine milk and meat production. The Scientific World Journal. Volume 2013, http://dx.doi.org/10.1155/2013/616098
Chung, H.R. 2014. Iodine and thyroid function. Ann Pediatr. Endocrinol. Metab. 2014;19:8- 12.
Cozzi, G., P. Prevedello, A. L. Stefani, A. Piron, B. Contiero, A. Lante, F. Gottardo, and E. Chevaux. 2011. Effect of dietary supplementation with different sources of selenium on growth response, selenium blood levels and meat quality of intensively finished Charolais young bulls. Animal, 5:1531–1538.
Depeint, F., W. R. Bruce, N. Shangari, R. Mehta, and P. J. O’Brien. 2006. Mitochondrial function and toxicity: Role of the B vitamin family on mitochondrial energy metabolism. Chemico-Biological Interactions, 163:94–112.
Huskisson, E., S. Maggini, and M. Ruf. 2007. The Role of Vitamins and Minerals in Energy Metabolism and Well-Being. The Journal of International Medical Research, 35: 277– 289.
Lenhinger. 1984. Bioquímica. Editorial Omega, México, D.F.
Millen, D.D., R. D. L. Pacheco, M. D. B. Arrigoni, M. L. Galyean, and J. T. Vasconcelos. 2009. A snapshot of management practices and nutritional recommendations used by feedlot nutritionists in Brazil. J. Anim. Sci. 87:3427–3439.
Mullur, R., Y. Liu, and G. A. Brent. 2013. Thyroid hormone regulation of metabolism. Physiol. Rev. 94:355–382.
NASEM. 2016. Nutrient Requirements of Beef Cattle (Eight revised edition). National Academies Press. Washington, D.C.
Nockels C.F., J. DeBonis, and J. Torrent. 1993. Stress Induction Affects Copper and Zinc Balance in Calves Fed Organic and Inorganic Copper and Zinc sources. J. Anim. Sci. Vo. 71:2539-2545.
NRC. 1958. Nutrient Requirements of Beef Cattle. Revised 1958. National Academy of Sciences-National Research Council. Washington, D.C.
NRC. 1984. Nutrient Requirements of Beef Cattle (sixth revised edition). National Academy Press, Washington, D.C.
NRC. 2000. Nutrient Requirements of Beef Cattle: Seventh Revised Edition: Update 2000. The National Academies Press, Washington, D.C.
Perry, T.W., W. M. Beeson, W. H. Smith, and T. Mohler. 1968. Value of zinc supplementation of natural rations for fattening beef cattle. J. Anim. Sci. 27:1674-1677.
Rossi, C.A.S., R. Compiani, G. Baldi, C.E.M. Bernardi, M. Muraro, J. P. Marden, and V. Dell’Orto. 2015. The effect of different selenium sources during the finishing phase on beef quality. Journal of Animal and Feed Sciences, 24:93–99.
Samuelson, K.L., M. E. Hubbert, M. L. Galyean, and C. A. Löest. 2016. Nutritional recommendations of feedlot consulting nutritionists: The 2015 New Mexico State and Texas Tech University survey. J. Anim. Sci. 94:2648–2663.
Sies, H. 1997. Oxidative stress: oxidants and antioxidants. .Experimental Physiology, 82:291-295.
Stangl, G.I., F. J. Schwarz, H. Mueller and M. Kirchgessner. 2000. Evaluation of the cobalt requirement of beef cattle based on vitamin B12, folate, homocysteine and methylmalonic acid. British Journal of Nutrition, 84:645-653.
Surai, F.P. 2006. Selenium in Nutrition and Health. Nottingham University Press. Nottingham, United Kingdom.
Vasconcelos, J. T. and M. L. Galyean. 2007. Nutritional recommendations of feedlot consulting nutritionists: The 2007 Texas Tech University survey. J. Anim. Sci. 85:2772–2781.
Veskoukis, A.S., M. Aristidis, M. Tsatsakis, and D. Kouretas. 2012. Dietary oxidative stress and antioxidant defense with an emphasis on plant extract administration. Cell Stress and Chaperones. 17:11–21.
Weiss, W.P. and M. Socha. 2005. Dietary manganese for dry and lactating Holstein cows. J. Dairy Sci. 88:2517-2523.
No hay comentarios:
Publicar un comentario